Jiao Xu1,
Xiao-hui Miao1,
Jin-xu Zheng2,
Ming Ding2,
Zhi-fang Zhuang1 ![]()
For correspondence:- Zhi-fang Zhuang Email: zhuangzfcdy@126.com Tel:+8651985336190
Received: 3 September 2016 Accepted: 10 February 2017 Published: 31 March 2017
Citation: Xu J, Miao X, Zheng J, Ding M, Zhuang Z. Therapeutic effects of Saikosapoin D on bleomycin-induced pulmonary fibrosis in mice via regulation of IL-33/ST2 pathway. Trop J Pharm Res 2017; 16(3):581-588 doi: 10.4314/tjpr.v16i3.12
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the therapeutic effects of saikosapoin D (SSD) on bleomycin (BLM)-induced pulmonary fibrosis (PF) in mice and its probable mechanisms.
Methods: PF mice were prepared by intraperitoneal (i.p.) injection of BLM (5 mg/kg). Twenty-four hours later, 72 mice in SSD group were administered SSD (1.8 mg/kg, ip). After 3, 7, 14 and 28 days of injection, the mice were sacrificed. Blood samples and lung tissues were collected from 6 mice in each group. The lung tissues were subjected to histological examination. In addition, ex
Results: Pathological results showed that SSD treatment alleviated alveolitis and lung fibrosis (p < 0.05) in lung tissues of PF mice at 14 and 28 days post-BLM injection. HYP and IL-13 levels of mice in SSD group were significantly lower than that in BLM group at days 14 and 28 post-BLM injection (p < 0.05). Levels of IL-4 and IFN-γ were significantly lower when compared with values in BLM group on day 28 (p < 0.05). Western blotting results revealed that ex
Conclusion: SSD exerts therapeutic effects on BLM-induced experimental PF in mice via regulation of IL-33/ST2 pathway
Introduction
Idiopathic pulmonary fibrosis (IPF) is a common and complex disease with high mortality and morbidity, and the median survival time is only 2 to 5 years [1,2]. Recent findings indicate that the mechanisms of its fibrotic effect involve transforming growth factor-β (TGF-β), Wnt ligands (secreted glycoproteins), toll-like receptor mediated signaling and type 2 immune responses, [3,4]. In addition, current clinical trials have evaluated various therapeutic approaches for IPF, such as dual endothelin receptor antagonist bosentan, imatinib, sildenafil, etanercept and interferon-c-1β, etc [5-7]. However, none of these ways attained the primary aims. Currently, the combination of corticosteroids and immunosuppressants is considered as common strategy for IPF treatment, however it would bring some serious adverse effects [8].
A novel signaling named IL-33/ST2 has been studied in many other fibrotic diseases, such as scleroderma, progressive systemic sclerosis and liver fibrosis [9,10]. ST2, an interleukin (IL)-1 receptor-related protein, is specifically expressed on Th2 cells, mast cells but not on Th1 cells, and has been recognized as a stable marker of Th2 cells [11]. IL-33, a novel member of IL-1 family, takes on its biologic activations by combining with its receptor protein ST2 [12].
Saikosapoin D (SSD) is an active constituent of Radix Bupleuri which is a famous herbal medicine in China with broad spectrum bioactivities. Previous reports indicated that SSD could be used to treat hepatic fibrosis and inflammatory diseases [13,14]. Thus, in the present investigation, we evaluated the potential therapeutic effects of SSD on bleomycin (BLM)-induced pulmonary fibrosis (PF) in mice, and the role of IL-33/ST2 pathway in this process.
Methods
Chemicals and reagents
Saikosapoin-D (SSD) was purchased from the Jiangxi Herbfine Science and Technology Co. Ltd (Jiangxi, China); chloral hydrate was obtained from the Jiangbin Hospital (Zhenjiang, China); Bleomycin (BLM) was purchased from the Nippon Kayaku Co., Ltd. (Tokyo, Japan); Hematoxylin and Eosin (H&E), Masson’s Trichrome, hydroxyproline (HYP) commercial kit, tissue RIPA buffer, primary antibodies for myeloid differentiation factor 88 (MyD88) and β-actin were purchased from the Wuhan Boster Biotech Co. (Wuhan, China); PVDF membranes and skimmed milk powder were obtained from the Millipore Co. Ltd. (Billerica, MA, America); primary tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6) monoclonal antibody was purchased from the Santa Cruz (Shanghai, China); commercial ELISA kits for IL-4, IL-13, IFN-γ and primary IL-33 monoclonal antibody were products of R&D. Inc. (Shanghai, China); primary ST2 polyclonal antibody was purchased from the Abcam Co. (Cambridge, UK); HRP-conjugated secondary antibody was purchased from the Cwbiotech. Co. (Shanghai, China).
Animals
SPF female C57BL/6 stain mice (6 weeks old, 18 ± 2 g) were purchased from the Laboratory Animal Center of Jiangsu University (Zhenjiang, China). All the animal experimental protocols were performed in accordance with the National Principles of Care and Use of Laboratory Animals [15] and were approved by the Ethics Committee of Changzhou Wujin People's Hospital (no. 2015a-4#A).
Animal model establishment and experimental protocols
A total of 72 mice were evenly divided into three groups (n = 24): control group (Cont.), BLM group and SSD group. After anaesthetized wtih chloral hydrate by intraperitoneal injection (0.01 mg/kg body weight, ip), mice in the BLM and SSD groups were injected with bleomycin (5 mg/kg, body weight, ip), while mice in control group were given equivalent volume of saline in place of bleomycin. Twenty-four hour after BLM injection, SSD mice were administered SSD (1.8 mg/kg body weight, ip). On day 3, 7, 14 and 28 after bleomycin injection, blood samples were collected using orbital blood sampling, and subsequently 6 mice of each group were sacrificed by cervical dislocation and the lung tissues were excised. The left lung tissues were fixed with 4 % paraformaldehyde, while the right lung tissues were frozen in liquid nitrogen for 10 min and then stored at -70 °C.
Histopathological investigations
The left lung tissues, fixed with 4 % paraformaldehyde, were paraffin embedded, and successively sliced at 4 μm thickness. The slices were stained with H&E to detect the degrees of inflammation and injury, while some were stained with Masson’s Trichrome to detect extent of fibros. According to methods established by Szapiel et al, score of 1, 2, 3, and 4 indicate grade 0, 1, 2, and 3 of alveolitis or pulmonary fibrosis was calculated [16].
Determination of HYP contents in lung tissues
The right lung tissues were used to determine the HYP contents according to the instruction on commercial HYP kits.
Western blotting assay
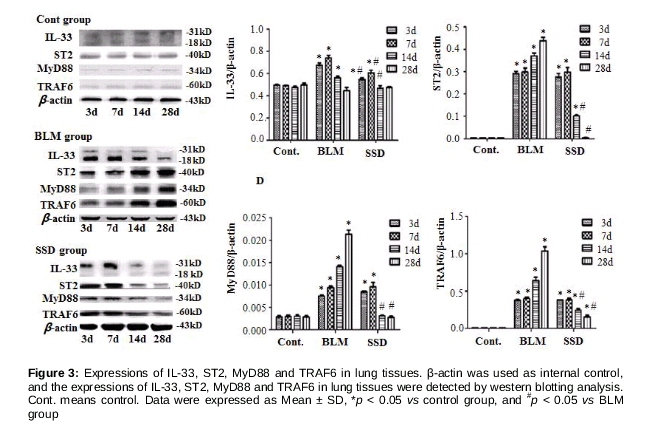
Lung tissues were homogenized and lysed with RIPA buffer and subsequently the total proteins were isolated. Then, equal amounts (30 μg) of protein were separated by SDS - PAGE and transferred to PVDF membranes. The procedures used followed the sequence: blocking with skimmed milk powder, incubation with primary polyclonal antibodies (including MyD88, TRAF6, IL-33 and ST2), and addition of HRP-conjugated secondary antibody. β-actin was used as internal control, and specific bands were visualized using ECL chemiluminescence kit (GE Healthcare, USA).
Determination of serum levels of IL-4, IL-13 and IFN-γ
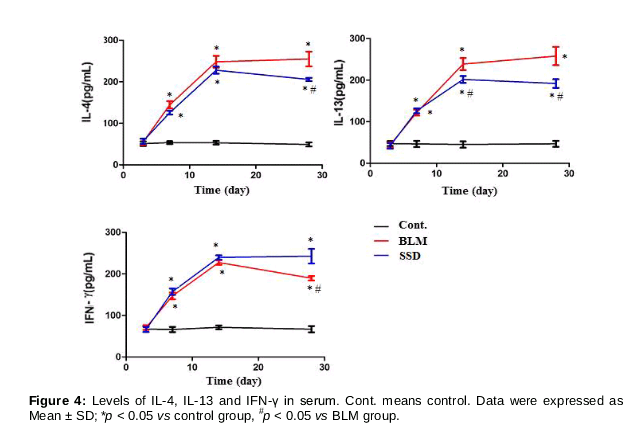
Blood samples were treated with heparin sodium and then stored at -70 °C. After blood sample were centrifuged, serum levels of IL-4, IL-13 and IFN-γ were measured by commercial ELISA kits according to the manufacturer’s instructions.
Statistical analysis
Data are presented as mean ± standard deviation (SD). Statistical significance of differences between groups was determined with SPSS 16.0 statistical software, using one-way analysis of variance (ANOVA) followed by LSD test (equal variances assumed) or Games-Howell test (Equal Variances not assumed). P < 0.05 was accepted as statistically significant.
Results
Pathological changes in lung tissues
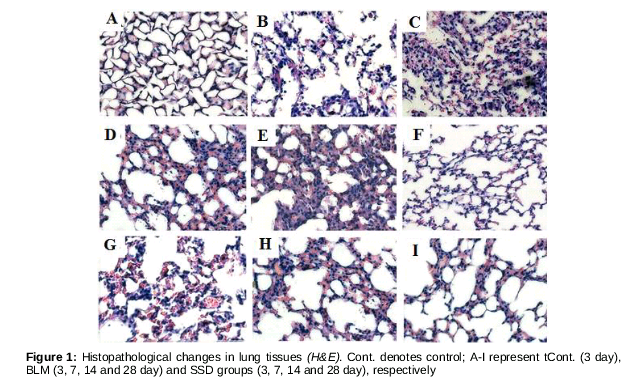
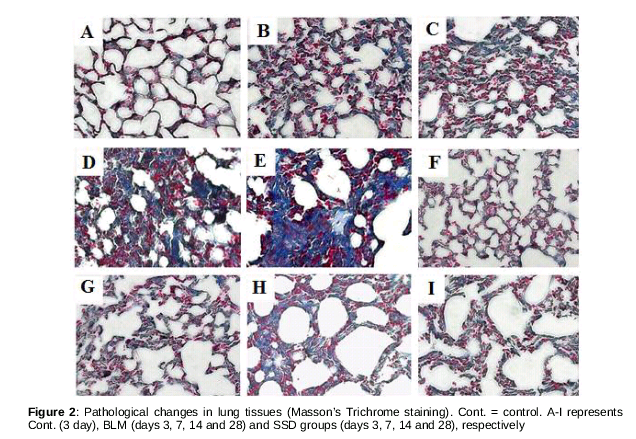
The architectures of lung tissues of mice in control group were normal without obvious inflammatory cells and fiber collagen proliferation (Figures 1&2). As can be seen from B - 1E, mild alveolitis was observed in BLM mice on day 3 after BLM injection; on day 7, the alveoli architectures was extensively broken, with evidence of high level of inflammation, and presence of inflammatory cells and erythrocytes in both septum and alveolus. Collagen fibers were few. The degree of inflammation on day 14 was milder, but the alveolus were collapsed and fused with extensive collagen fiber proliferation. On day 28, mild inflammation was observed, but the architectures of alveolus were blurred, with wild broad-band collagen fiber, and diffuse lung fibrosis. The degree of alveolitis in SSD mice on day 3 was not different from that of mice in BLM group. Interestingly, alveolitis in SSD-treated mice on day 7 was less than that of the BLM-treated mice. On day 28, the extent of alveolitis in SSD mice was less than that in BLM mice, with fewer collagen fibers. However broken and disordered architectures was still evident ().
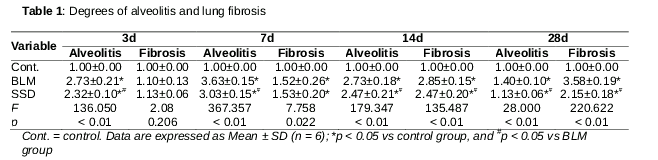
There were significant differences on days 3, 7, 14 and 28 in grades of alveolitis in BLM and SSD groups compared to control group (p < 0.05, ). However, the alveolitis of SSD-treated mice were milder than that of BLM mice at all time points, with significant differences on day 28 (p < 0.05, ). Compared to control group, fibrosis grades of both BLM and SSD mice significantly different on days 7, 14 and 28 (p < 0.05, ).
HYP levels in lung tissues
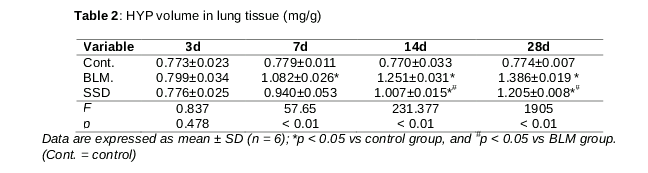
The concentrations of HYP in lung tissues from both BLM and SSD groups increased and reached the peak on day 28. There were significant differences on days 7, 14 and 28 in BLM group, compared to control group (p < 0.05, ).); while the levels of HYP in SSD group were less than that in BLM group from day 7, and became significantly different on days 14 and 28 (p < 0.05, ).
Expressions of IL-33, ST2, MyD88 and TRAF6 in lung tissues
Compared to control group, the protein levels of ST2, MyD88 and TRAF6 in BLM mice increased as from day 3, with significant increases on days 3, 7, 14 and 28 (p < 0.05); while the protein levels in SSD mice increased initially, peaked on day 7, and subsequently decreased. Significant differences ST2 between this group and control were obtained on days 3, 7 and 14, and on days14 and 28 relative to the BLM group (p < 0.05). There were significant differences in MyD88 between the SSD group and control days 3 and 7, and between the SSD and BLM groups on days14 and 28 (p < 0.05). Significant differences in TRAF6 between the SSD group and controls were obtained at all times points control group (p < 0.05), while on days 14 and 28 versus BLM group (p < 0.05) (). The IL-33 level increased firstly then decreased gradually, with its peak on day 7. Significant differences were observed on days 3, 7 and 14 in BLM mice, relative to control and SSD mice (p < 0.05); while compared to control group, there were significant differences on days 14 and 28 in SSD group (p < 0.05, ).
Serum levels of IL-4, IL-13 and IFN-γ
The concentrations of IL-4, IL-13 increased from day 7, and became significantly higher on days 7, 14 and 28 relative to control group (p < 0.05); there were significant differences in IL-4 between the BLM and SSD groups on day 28 (p < 0.05), while for the same groups, IL-13 levels differed significantly on days 14 and 28 (p < 0.05) (). Values of IFN-γ in BLM and SSD groups rose from day 7 and peaked on day 14, with the same significant differences on days 7, 14 and 28 relative to control group (p < 0.05); meanwhile, at every time point, the levels of IFN-γ in SSD group were higher than that in BLM group, and significant differences were obtained on day 28 between two groups (p < 0.05) ().
Discussion
Bleomycin (BLM), an anti-neoplastic antibiotic is a derivative of Streptomyces verticillus with severe side-effects as inflammation and fibrosis specific to the lung [17]. To the best of our knowledge, no suitable treatment exists for lung fibrosis currently. Animal models are important the investigation of pathological mechanisms and preclinical evaluation of novel therapies. Therefore, bleomycin-elicited injury in small rodents have been become the traditional and most commonly adopted lung fibrosis model [18]. The results showed that when the lung exposed to bleomycin, there were damages to the lung tissues, accompanying some epithelial or endothelial cells over-apoptosis passively respond to the injury. In addition, the SSD could be used to effectively ameliorate the damages induced by bleomycin.
IL-33, as an ‘alarmin’, was secreted from the injured cells. This study further detected MyD88 and TRAF6, the down-stream molecules of ST2, which can ultimately lead to the activation of NF - κB and MAP kinases involved in the control of cellular proliferation and apoptosis [12]. Expressions of protein MyD88 and protein began to increase from day3 post-model, and attained maximum levels on day 28, with similar trends for protein ST2, IL-4 and IL-13. IL-4 and IL-13 are two major cytokines of Th2 cells, which can involve fibrogenesis by up-regulating proteins associated with wound healing, specifically arginase, collagens, matrix metalloproteinases (MMPs), and tissue inhibitors of MMP [19] or via recruiting M2 cells. In addition, they also could facilitate excessive tissue repair and fibrosis [20]. Therefore, from our results, it is speculated that IL-33 secreted from the dying or apoptosis cells via its receptor ST2 (a stable maker of Th2 cells). And IL-33 also amplified Th2-type responses and involved pulmonary fibrosis process. Whereas, compared to BLM group, the expressions of IL-33 in lung tissues decreased from the day 3, and still remained the same trends. Interestingly, levels of ST2, MyD88 and TRAF6 in SSD group increased firstly then decreased gradually, peaking on day 7.
The dynamic changes in IL-4, IL-13 and IFN-γ further demonstrated that IL-33/ST2 signaling pathway was activated and a Th1/Th2 imbalance existed in the pulmonary fibrosis process. Herein, it revealed that corticosteroids, as conventional ‘anti-inflammatory’ therapy for fibrosis diseases, can disturb the expression of IL-33/ST2 signaling pathway and the balances of Th1 and Th2 cytokines. Nevertheless, more recent studies have adopted other methods to explore the role of IL-33/ST2 signaling pathway in many disorders. Mingcai Li et al demonstrated that blockage of IL-33/ST2 signaling pathway by anti-IL-33 antibody can inhibit airway inflammation in a murine model of allergic asthma [21]. Hiroko Hayakawa et al showed that the antagonistic effects of soluble ST2 on IL-33 signaling using a murine thymoma EL-4 cells stably expressing ST2L and a murine model of asthma [22]. The results of present study demonstrated that the antibody or the soluble protein and corticosteroids exhibited impact on the IL-33/ST2 signaling pathway. Thus, the further investigations need to be done to find out the effective blocking agents for reducing the degree of fibrosis. To the best of our knowledge, the present study is the first to report that IL-33/ST2 signaling exist in a murine model of bleomycin-induced pulmonary fibrosis. It is also the first study that to report that Saikosapoin-D has some degree of interference in IL-33/ST2 signaling pathway, compared to other studies that found that IL-33/ST2 signaling pathway was interfered by anti-IL-33 antibody [21] or soluble ST2 [22], or administration of IL-33 [23].
Conclusion
These findings demonstrate that SSD is a potential candidate for treating IPF via down-regulation of IL-33/ST2 signaling. In addition, the results also suggest that IL-33/ST2 signaling pathway is a potential drug target for treating IPF.
Declarations
Acknowledgement
References
Archives


News Updates